※勢いで書きました。修正・加筆する可能性があります。ご質問・ご指摘があれば、遠慮なくメールにてご連絡ください!
0. はじめに
生命とは、高分子 (DNAやタンパク質) や脂質膜などのソフトマターと呼ばれる分子群が、代謝と呼ばれる化学反応ネットワークを介して連携することで、自己生産や進化などといった自然界の他の物質系には見られない特異的な振る舞いを示すシステムです。私のサイエンスは、この生命という特異な物質の存在様式の出現を可能とする原理は何なのかを、物理学を基軸とした物質科学の立場からボトムアップに問うことにあります。
-
図1:脂質膜ベシクル
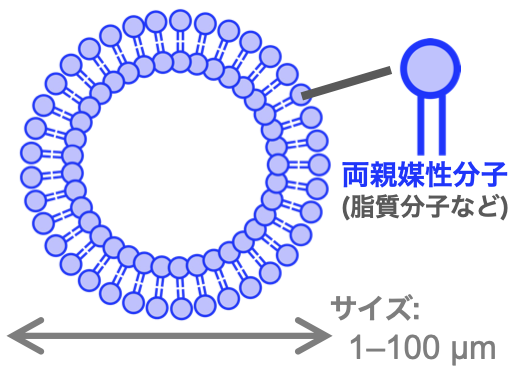
中でも私は「自己生産して増える」という生命の基本性質にまず興味を持ちました。そこで私は「細胞」の字義の通り、まずミクロな脂質膜コンパートメントであるベシクル(図1)に狙いを定めました。そしてこの「ガワだけの空っぽの細胞」に相当するベシクルが変形・分裂するための膜の物理と、ベシクルが成長するための化学反応系とを組み合わせます。これにより、単純な分子系から、実際に自己生産などの生命の基本性質を極めてシンプルに再現するモデル実験系「(人工)ミニマルセル」を作ってみるというアプローチで、物質と生命を繋ぐソフトマターの組織化原理の解明に挑んできました。
また九州大に移ってからは、これまで私のターゲットだった細胞の「ガワ(膜)」だけでなく、細胞の「中身(細胞質)」の動的な、極めて興味深い物理的性質を解明する取り組みにも参加しています。将来的には、私はガワも中身もひっくるめた、細胞全体に対して物性物理学の立場からアプローチしていきたい。そしていつの日か、それ自体は生きていない物質がどのような原理に従って「生きている」という状態・システムへと組織化されるのか、すなわち「物質に生命が宿るとはどういうことか」という究極の問いに対し、自分なりに納得できる物理学的世界観 (物理学からの世界の見方) を、何か一つでも生み出すことが出来ればなあ、と夢見ています。
1. ボトムアップ人工細胞研究(人工ミニマルセル)
生命を理解する上での最大の障壁は、システムの圧倒的な複雑さにあります。その構成要素も膨大です。しかし物理学の観点で生命を理解したい時、我々が期待するのは、生命の特異性をそうしたシステムの構成要素の特殊性に帰着させる類の答えというよりも、むしろシステムの構造や組織化の普遍的形式を見出して提示する類の答えなのではないでしょうか。我々はセントラルドグマと呼ばれる、DNA-RNA-タンパク質系から成るシステムしか知りません。しかしそれを通して見出された性質が、本当に生命という物質の存在様式に“普遍的”なのか、それとも生命をDNAなどの特定分子で構成する場合に限って現れる“特殊な”条件に過ぎないのか、1つの系しか知らない我々に区別は困難だろう。極端な考えではありますが、学部・修士時代の私は「生命と複雑系」(田中博, 培風館 2002 (残念ながら絶版済))という本の影響を大いに受けたことで、そんなことばっかり考えていました。
そこで我々は、「できるだけ我々の知る生命とは違う分子や反応を使ってみる」という縛りを自らに課し、その上で生命の代謝や自己生産などの振る舞いを最小限に実現するようなモデル実験系(人工ミニマルセル)を創出できないものかと取り組みました。生命の本質的な挙動を人工のモデル系でシンプルに再現し、天然系と人工系、2つの共通部分・異なっている部分を比較すれば、物質科学ベースで生命を問えるのではないか?
ここで、自己生産に必要不可欠な膜コンパートメントの変形・分裂の物理については当時私の指導教員だった今井正幸先生(東北大)がエキスパートでした。しかし実際に実験系を構築するに当たって、物理学の範疇の技術では限界がありました。そこで学部4年生の頃から超分子化学者のPeter
Walde先生(スイスETH)と共同研究させてもらうことで、膜コンパートメントに重合反応や(本当は使いたくなかったのですが)酵素反応を組み合わせる方法を検討することができました。
-
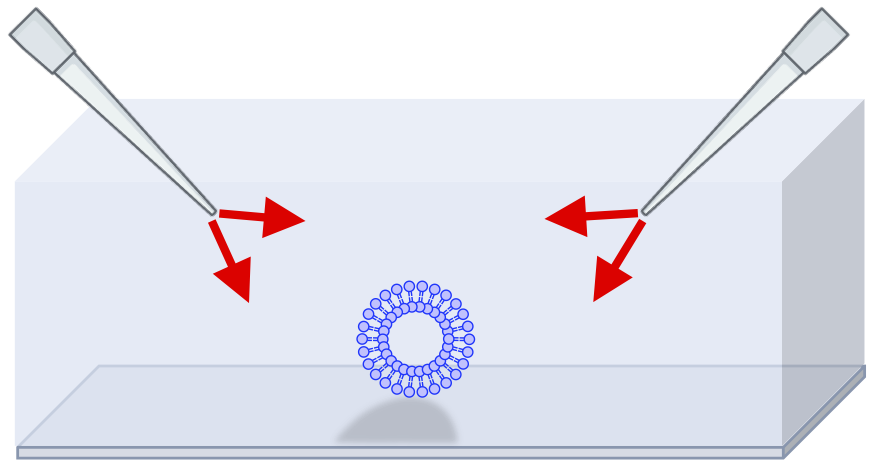
図2:人工ミニマルセルの実験系
-
図3: 人工ミニマルセルの代謝系
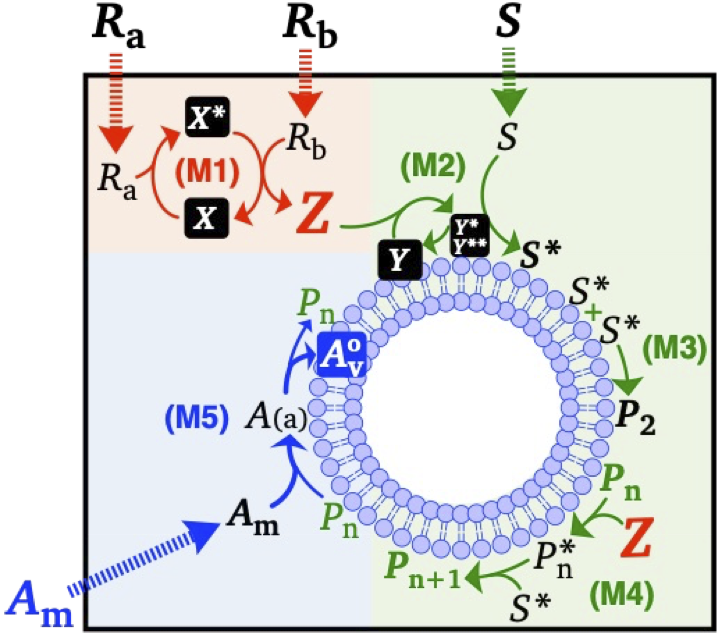
図2がその実験系の様子です。材料分子や触媒分子・反応環境を整えた培養液のような環境に、変形・分裂しやすいように組成を調整した脂質膜ベシクルが沈んでいます。そこに両サイドからそれぞれ異なる材料分子や膜分子を供給し、2つの水流の間にベシクルをトラップしてやります。これにより、ベシクル近傍に定常的にフレッシュな材料分子の流入、および廃棄物の流出が促されます
(“散逸構造”を意識しました)。
こうした環境設定にて、図3のような化学反応系がベシクルの表面に生じます。3色の区分は反応系の役割の違いを表していて、赤色では酵素反応によって2つの材料分子 (酸素とグルコース,
RaとRb) から下流の反応系を駆動するためのエネルギー分子 (過酸化水素, Z) を生成します。緑色ではエネルギー分子 (Z) と別の材料分子 (アニリン, S) から、ベシクルを成長させる触媒として機能する高分子 (PANI-ES, Pn) を膜表面に合成します。最後に青色では、追加の膜分子 (Am) のベシクル膜への取り込みが、膜表面に合成されている触媒高分子(Pn) によって促進されます。
これらの結果、3ユニットからなる化学反応系(図3)が定常的な材料供給に伴い誘起することで、ベシクル表面が反応場となって触媒高分子を合成し、今度はその触媒高分子が反応場であるベシクル膜の成長を促進するという、ベシクルと高分子の相互触媒系が形成されます (Kauffmanの “Autocatalytic Set”を意識しました)。これによってベシクル・高分子系は急速に成長していくのですが、ただ大きくなって終わりではなく、脂質膜ベシクルの成分を膜弾性理論に基づいて調整することで、ある程度ベシクルが成長した後に自発的に変形・分裂するように誘導することができます。最終的に、図4や動画のように、我々は材料分子供給下で第4–5世代程度まで自発的に自己生産する人工ミニマルセル系を単純な分子系から創出することに成功しました (この研究で私は博士号をもらいました)。
-
図4: 自己生産する人工ミニマルセル

人工ミニマルセル系の良いところは、系の全貌が単純で、系の全体像を数理的に記述することが(比較的)容易なことです。例えば、ベシクルの成長については水溶液中での供給膜分子の化学ポテンシャル勾配によって、化学反応系のキネティクスについては一つ一つの反応速度式を書き下すことによって、またそれらと連携したベシクルの自発的な変形・分裂については膜の曲率ストレス(自発曲率)を考慮する弾性モデルによって記述することができます。
こうしたモデル実験系としての数理的な記述可能性は、物質と生命を繋ぐ物理的原理の探究の上で必要不可欠であり、現在も取り組みを継続しています。例えば最近は、理論物理や生命科学の若手研究者たちと共同して「生命と非生命の本質的な違いを理解しよう」という取り組みを始めています
(参考:学変B「生命-非生命転移」)。また「自己生産できるなら、次は進化できるのか?」というコメントは、どこかしらで研究発表するたびにほぼ毎回投げかけられています。
究極的には、それ自身は生きていないソフトマター分子系がどのように組織化・秩序化することで「生きている」という状態のシステムが出現するのか、その理解や、物理学的世界観の構築のための道具として使えるといいなあ、と夢見ています。
関連する論文:
Kurisu et al., "Synthesising a minimal cell with artificial metabolic pathways",
Communications Chemistry 6, 56 (2023).
Kurisu et al., "Reproduction of vesicles coupled with a vesicle surface-confined enzymatic
polymerisation", Communications Chemistry 2, 117 (2019).
Kurisu et al., "Vesicle-surface-templated catalytic polymers drive
differential growth in synthetic minimal cell variants" (submitted).
2. 汎用的なマイクロ増殖容器~「中身」を入れて自己生産させたい~
現在、脂質膜ベシクル(図1) の内部に機能性分子や化学反応を格納することで、細胞サイズの高機能な分子システムを創出しようという試みが世界中で行われています (人工細胞などと呼ばれます)。しかし多くの場合、これらの取り組みでは中身として搭載する分子や化学反応の開発には注力する一方で、ベシクル自体はただの容器として扱われ、あまり注力されている印象はありません。しかし、もし内部にモノを格納した状態で簡単にかつ大量に分裂増殖できる容器があれば、現在盛んに研究されている高機能な中身をそのままに容器の部分だけを置換してアップグレードすることで、様々な能力を持つ人工細胞たちに対して一挙に増殖能力も付与することができるかもしれません。
-
図5: 分裂増殖系の駆動環境
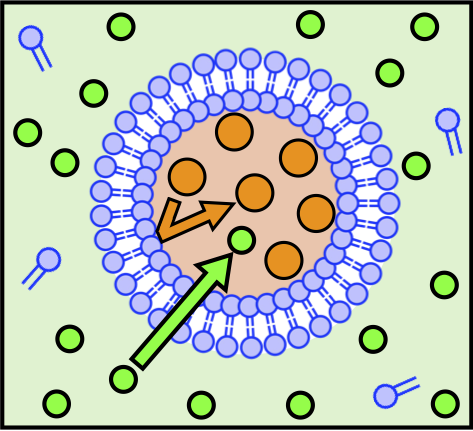
そんな期待が持てそうな分裂増殖系を偶然、私は発見しました。分裂条件は非常にシンプルです。まず2種類の膜分子 (AOTとコレステロール) で構成されたベシクルに対し、図5 のように、外側に膜を透過しやすい分子(緑色, 例えば単糖分子)、内側に膜を透過しにくい分子(橙色, 例えば二糖分子や高分子) の水溶液を配置します。人工細胞を設計しようとする時、ほとんどの場合でベシクル内部に分子量の大きな貨物分子を格納するので、それと同様のセットアップになっています。ここに、外側の水溶液に対して追加でベシクルの構成分子(青色, AOT) を溶かしておくことで、自発的かつ再帰的に出芽型分裂が駆動されます。
具体的なメカニズムとしては、膜に対して透過速度が異なる分子がベシクル内外に非対称に分布していることで、球形膜を外に押し広げるような浸透圧がじわじわと働き続けます。その浸透圧が生み出す膜の張力ストレスを緩和するために、ベシクルは外側の水溶液に溶けている追加の膜分子を膜に取り込みますが、この時それらの膜分子は脂質二分子膜構造の外膜から挿入されていくことになります。これにより、外膜・内膜間に生じる膜の非対称性が二分子膜に曲率ストレスを蓄積させ、最終的に膜の出芽型変形へと至るのだと考察しています。
-
図6: 分裂増殖ベシクルの様子

-
図7: 親半径(横軸)に対する子半径(縦軸)
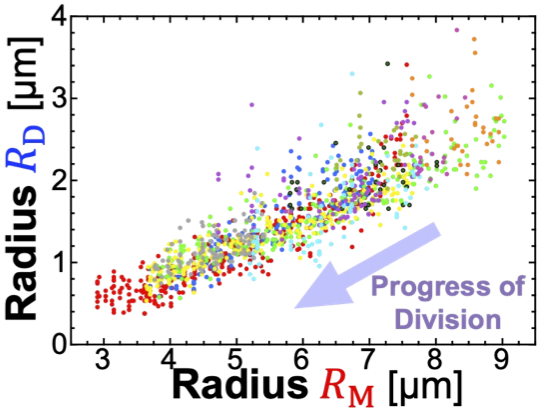
図6と動画が分裂増殖の様子です。個体差はありますが、大体一つの親ベシクルから30–300個もの子ベシクルが、自発的な出芽型分裂により次々と産まれ続けます。沢山の子ベシクルがイクラのように見えたのと、親がまるで卵を産んでいるように見えたことから、この新しく発見した現象を“Osmotic Spawning Vesicle”(=浸透圧産卵ベシクル?)と名付けました。
このOSベシクルにおける実用上の重要な性質が、写真を見ても何となく分かりますが、産まれてくる子ベシクルのサイズがランダムではなく、ある程度揃っていることです。図7 に親の半径に対する子の半径をプロットしたものを10測定分重ねて表示していますが、確かに親の初期サイズや分裂の経過時間に関わらず、「同じサイズの親からは大体同じサイズの子が産まれる」というサイズ制御性があることが分かっており、このことは膜弾性モデルからも理論的に説明可能です。
また、実際の細胞では極めて複雑なタンパク質機構を用いて細胞分裂という生物学的機能を実現しているのに対し、OSベシクルでの分裂増殖は化学反応や機能性分子を必要としない、純粋に物理的な系です。そのため、仮にこのベシクル系の内部に人工細胞のための様々な化学反応系や機能性分子を格納したとしても、それら人工細胞の中核機能に干渉したり・されたりすることなく、独立に分裂の制御ができる見込みがあります。膜コンパートメントをベースとした様々な人工細胞系に対し、将来的に単純かつ一挙に増殖機能を実装する仕組みとなり得ると期待しています。
また物質と生命を繋ぐソフトマターの組織化原理という視点に立ち返ると、わずか2成分の膜が、これほど単純な条件下で自発的に分裂増殖を行い出すという事実は衝撃です。この極めて原始的な分裂増殖系の成立条件や射程を探ることで、「自己生産する細胞」という構造が単純な分子系から出現し得るための物理的要件を明らかに出来るかもしれません。
現時点では、ベシクル膜が出芽型変形を行うのか、それとも変形せずに球形を保ち続けるのかを決定的に左右する要因として、膜の熱ゆらぎの挙動が関係していると私は睨んでいます。そして膜の熱ゆらぎには、膜の周囲を取り巻く流体の粘弾性的性質が大きく影響します。現在のOSベシクル系はまだプロトタイプで、中身としては主に二糖水溶液を用いてきました。しかし実際の細胞の中身には、より粘度が大きく、弾性的な性質も併せ持つ細胞質や、大小様々な細胞内小器官が詰まっています。
単なる膜としてだけではなく、その内部に格納された流体やコロイドとの相互作用系として理解していくことで、最終的には、自己生産するミニマルな細胞構造の出現が許容される物理パラメータ空間の相図、いわばソフトマターにおける「生命出現許容領域 (life-permitting landscape)」のようなものを描き出せないものかなあ、と夢見ています。
関連する論文:
Kurisu et al., "Osmotic spawning vesicle",
Soft Matter20, 8976-8989 (2024).
栗栖実,「出芽型分裂で増殖する脂質膜ベシクルの開発」,
生物物理 65, 248-251 (2025).
3. 細胞の中身だけを取り出して、生命力(?)を測定する
私は学生時代から頑なに実際の生命には触れてきませんでしたが、九州大に移ってからはその縛りを解き、最近は博士課程の冨山君のお手伝い役として、この研究に携わっています。
生物は生きている時は何となく活動的で、死ぬと静止する印象を私たちは持っています。この“印象”を物理学の言葉で上手く言い表せないものでしょうか?関連するかもしれない現象として、ゼリー飲料は振ることで液体のようになりますが、しばらく置いておくと流動性を失い、徐々に硬くなって固体のようになります。実は細胞の中でも同様のことが起きていると考えられています。
本来、細胞の中身 (細胞質)は非常に物質濃度が高く、分子混雑している状態です。そのため、生きている細胞から中身だけを取り出して放置していると、自然と流動性を失っていき、硬くなり、やがて固化 (ゲル化) してしまいます。にも関わらず、生きている細胞の中に在る限り、細胞質は活動的なままであり、どうにかして流動性を保っていて固化しません。
その詳しい機構はいまだ分かっていませんが、細胞質が生きた細胞の内部に在るときには、常にエネルギー分子 (ATPなど) を消費しながら大規模な化学反応系
(代謝) が駆動されています。現時点での仮説としては、この代謝によって細胞質が激しくかき混ぜられ続けられることで、流動的な状態が維持されている一方で、そこから中身を取り出して放置してしまうと、細胞質がエネルギー切れになって自身をかき混ぜられなくなり、それゆえ自然に固化してゆく作用にもはや抗えなくなってしまうというシナリオなのではないか、と考えられています。
-
図8: 細胞抽出物の力学測定
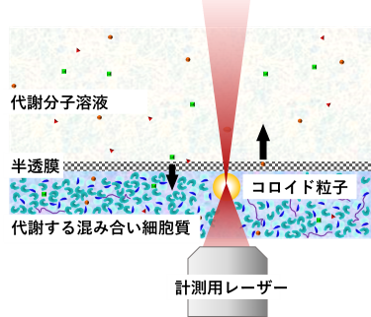
この細胞が持っている流動性維持(生命力維持?) 機構を支える実体を解明すべく、我々は図8 のように、生きている細胞から抽出した中身を半透膜の中に閉じ込める実験系を用いています。抽出した細胞質には、半透膜を介して外部から生理活性物質(ATPなど)を供給するとともに、代謝によって生じた老廃物は外部に排出されます。これにより、細胞から中身を取り出した状態であっても長時間にわたって代謝活動を持続させることができます。そして取り出した細胞質の中にコロイド粒子を置き、粒子がその中でゆれ動く様子を追跡することで、我々は現在、細胞質のかき混ぜる力によって生じる非平衡ゆらぎの強さを測定中です。
究極的には、細胞質の組成操作も行いながら、様々な細胞種について代謝活性に対する流動化作用を測定していくことで、「生きている」状態のあらゆる細胞の中身に共通して働いている普遍的な原理を明らかにできればなあ、と夢見ています。
関連する情報: 九州大・複雑生命物性研究室(水野研) ホームページ